赵杨
中国科学院分子植物科学卓越创新中心
研究员 博士生导师
| 剁开一粒黄土,半粒在喊渴,半粒在喊饿”。“根冠比调控”一直是植物抗逆领域关注的核心科学问题,为什么干旱能抑制植物地上部分的生长,却会促进根系的生长?破解植物抗逆机制,培养抗逆、节水、高产、高质量的农作物,是每一位逆境研究者的梦想。 |

3'46"
复水之后它们就又存活,这是一种非常强大的力量。在我国的神话传说里面有一种九死还魂草,它其实就是复苏植物——卷柏。而卷柏的复苏,它就像怒放的生命一样,卷柏它的生命力极为顽强。

11'56"
我们发现了这么一个调控,它就像打通了植物体内蔗糖的一个物流的转运。让叶片合成的这种蔗糖快速地转运到需要它的地方,就像根、芽和果实中,从整体上提高了植物的抗旱和生长。
14'44"
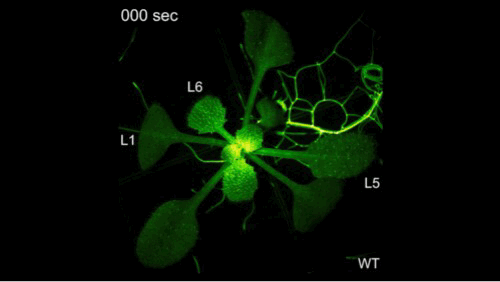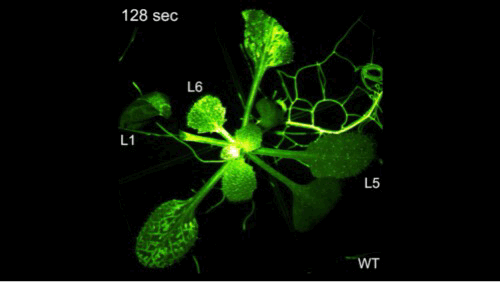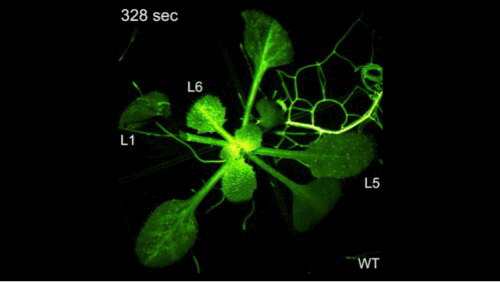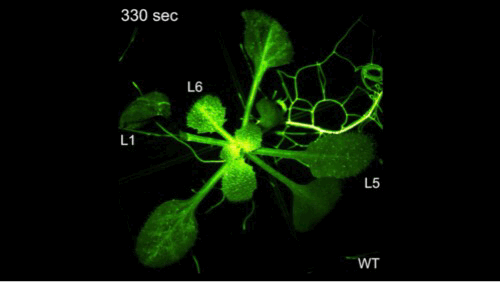
钙信号它是非常快的,但是有多快呢?大家看到了,剪掉一片叶子之后,钙信号瞬间爆发并且传遍植物的全身。植物它并不是不能感知,它只是不会跑而已。
我出生和成长在黄土高原,在我的记忆里,干旱就是刻骨铭心的缺水。每年的冬春季节村庄断水,我就跟着爷爷、叔叔走十里山路,到别的村庄去拉水,水和粮食一样珍贵,就像一位诗人所说,“剁开一粒黄土,半粒在喊渴,半粒在喊饿。”这种记忆是刻骨铭心的,直到现在我还舍不得浪费水。

黄土高原是我国华夏文明的中心,也是起源地,但是为什么这一地区干旱频发呢?我们可以从地理书中找到答案。我国的东部是典型的大陆季风气候,东南的季风带来水汽,但在我国的西北地区它是非季风区,常年干旱。
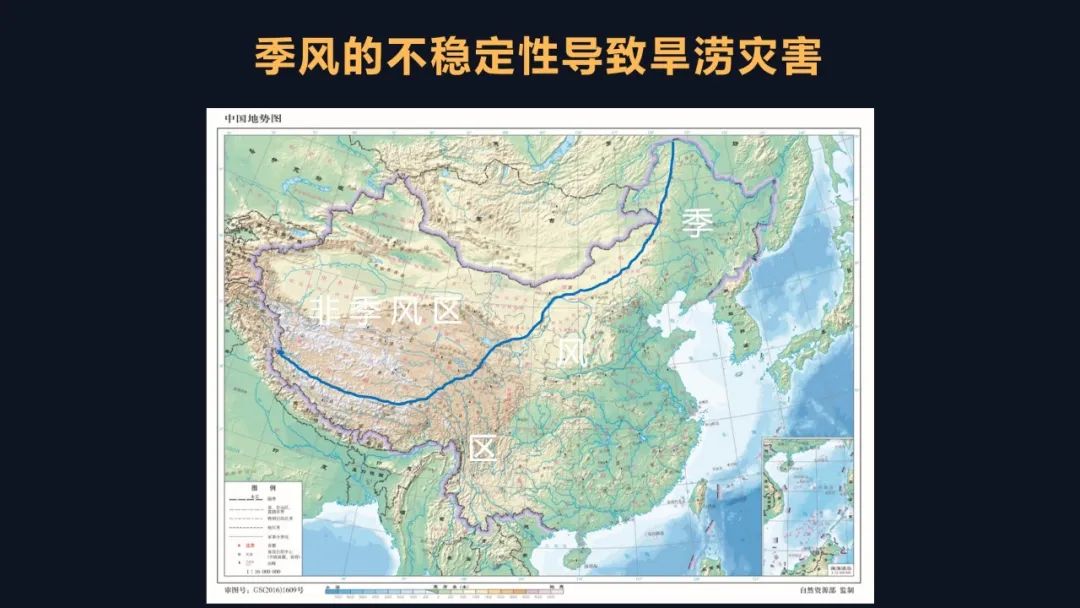
中华五千年的文明也是一个水旱灾害的防御史。《史记》记载了前三千年的历史,司马迁描述道,“六岁穰,六岁旱,十二岁一大饥。”这是什么意思呢?三千年前平均两年一次干旱,十二年一次大饥荒。在接下来的两千年里,大约发生了1300次大旱。也就是五千年的历史里面,一直是平均两年一次。干旱如此频繁,它会带来如何严重的后果呢?在《国语》中记载“伊洛竭而夏亡”。也就是河南陕西一带的伊河、洛河干涸了,导致了夏朝的灭亡。那在之后的朝代里,例如明朝末年,连年大旱,农民起义,明朝灭亡。

新中国成立之后,我们国家兴修水利,人民丰衣足食,然而干旱真的离我们远去了吗?其实不然,我们可以预见到未来依然缺水。在接下来的三十年里,人口将要进一步增长,需要的粮食也进一步增长。然而我们的耕地面积有限,而淡水资源是在逐渐地匮乏,也就是说粮食危机依然存在,因此我们才要研究植物如何应对干旱,以及如何培育抗逆稳产的作物。
为什么水对我们的生命如此重要呢?这是因为我们地球的生命起源于水。人体中有65~70%是水分,植物体中有80~95%,当我们失水2%开始感到口渴,4%的时候皮肤失去弹性,8% 我们的精神异常,10% 我们危及生命,20% 死亡。

当动物感到口渴之后,它可以去寻找水源,但是植物它由于固着生长无法移动。在长期的进化中,植物它也就进化出来各种各样抗旱的策略,像花粉散播、种子存储,以及旱区长期生长的植物,它们都要经历长时间的干旱胁迫,它们进化出了一种“干而不死”的策略。

这些细胞它如何在脱水的环境里面存活呢?我们可以看到在干旱下,它们会积累大量的高度亲水的无序蛋白。这些蛋白可以保护我们生命的大分子,就像蛋白质 核酸等等。它保护这些大分子不变性,干旱下它们会进一步的脱水收缩,细胞变成了一个水凝胶的状态。生命似乎按了暂停键,进入了休眠状态。

复水之后它们就又存活,这是一种非常强大的力量。在我国的神话传说里面有一种九死还魂草,它其实就是复苏植物——卷柏。而卷柏的复苏,它就像怒放的生命一样,卷柏它的生命力极为顽强。它在3亿年前进化出来,然后至今还生活在我们旱区的石缝中,是一种先驱植物。

但是它的缺陷是什么呢?它长得极慢。在长期的进化中,我们植物还形成了一种应对干旱的更普遍的策略避免脱水。植物进化出叶片,它可以进行光合作用,将光能转化为化学能。植物还进化出来根,可以从土壤中吸取水分,在叶片上有一个个小孔就像鼻子一样,我们叫做气孔。它可以吸入二氧化碳,放出氧气和水。

在干旱的环境下,植物关闭气孔,减少水分的丧失。在沙漠植物里面,还进化出来各种各样抗旱的策略,例如仙人掌,它把叶片进化出像针刺一样。猴面包树它可以用树干储水,骆驼刺有非常深的根系。生石花将自己封闭,当厄尔尼诺带来沙漠的大雨,潜藏的种子,它可以快速地萌发生长,开花结果,完成一个生命周期。在短短的几天里面,一个生命禁区就焕发了勃勃的生机。

它们似乎懂得深挖洞、广积粮,高筑墙、缓称王的策略。进化的过程里面,五亿年前植物开始登陆,登陆之后,在干旱的环境下倔强地生存和繁衍,将我们一个原始荒凉的地球大陆,变得如今生机勃勃。在这个过程里面有一个抗旱的激素,ABA又叫脱落酸,它起了核心的作用。脱落酸它是如何被发现的呢?在上世纪六十年代,来自美国加州大学 英国威尔士大学,和新西兰科学与工业管理局,三个实验室他们分离出一种小分子。这种小分子,它可以促进桦树芽的休眠,帮助它度过严寒和干旱,还可以调控棉铃的脱落,黄羽扇豆花和果实的脱落。科学家将它命名为休眠素和脱落素II。在之后,发现这个小分子实际上是同一个物质。在1967年,在第六届的植物生长物质会议上,大家协商了之后把它命名为脱落酸。

脱落酸发现之后,科学家就开始研究它的合成信号功能,但是脱落酸的受体直到2009年才被发现,它是我们植物中,五大激素里面最后发现的受体。因为脱落酸的受体家族成员非常庞大,传统的方法找不到。加州大学有一位科学家Sean Cutler,他就利用一种新的策略,他先寻找脱落酸的类似物,再利用类似物去寻找受体,最终他成功地找到了这个受体。他从博士期间就已经开始脱落酸的研究,前后用了近20年,最终才找到脱落酸的受体,解决了一个世纪难题。与此同时,在德国Erwin Grill实验室,他们用生化的方法也找到了脱落酸的受体,他们共同的发现,在2009年被《科学》期刊,评为十大科学进展之一,并被广泛应用于作物的抗逆遗传改良。

脱落酸它有什么样的功能呢?脱落酸的功能,我们可以看一看正常的植物是怎么去生长。在适宜的环境下,种子要萌发生长,开花结果,但是干旱它会打断这个环节,让植物死亡。而脱落酸它可以促进种子的休眠,抑制萌发、抑制生长,让气孔关闭,植物虽然长得小,但它很强壮,可以在干旱下存活。脱落酸还有一个重要的功能,就是促进休眠,让种子具有非常长的寿命,可以看成是一个长寿的激素。在我国和日本多地发现了一种,千年甚至两千年的古莲子,在经过精心的培育之后,它们依然可以开花结果。脱落酸在这中间就起到重要的作用。

它的功能可以总结为忍耐和等待,忍耐暂时的逆境,等待顺境的来临。由于脱落酸的特殊性,我们在植物抗逆中就有一个难题,生长和抗逆不可兼得。当我们关闭气孔,减少水分的丧失,这时候就带来一个问题,二氧化碳的进入也减少,光合减少,光合减少之后,根系就不能正常的生长,没有根系,我们就无法获得水分。因此植物采取了这种忍耐和等待的策略,最终就是进入一个闭关锁国的状态,坐吃山空。
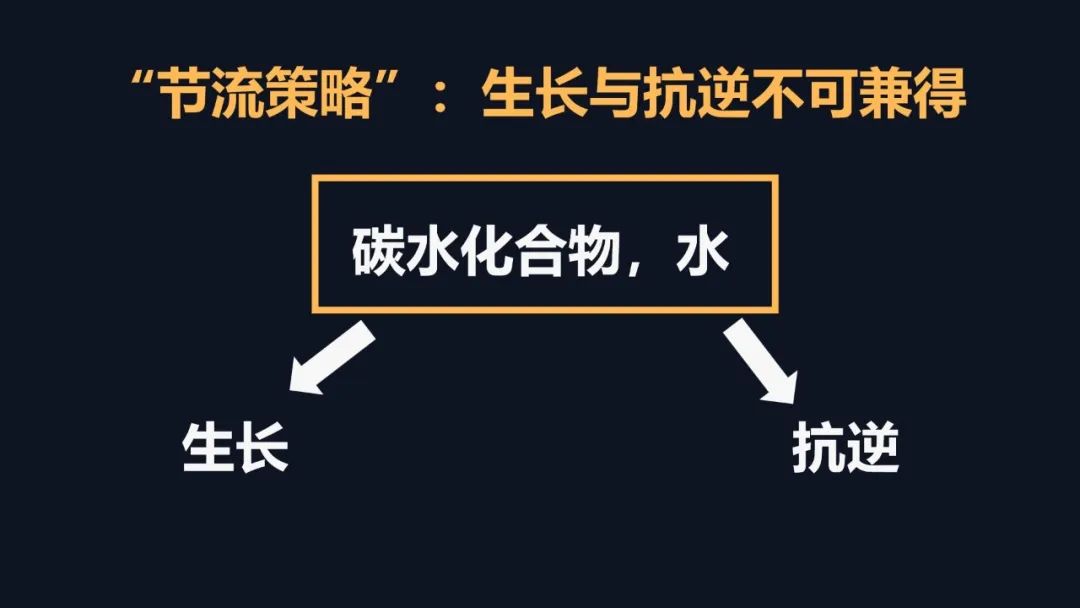
我之前的研究,我还陷入过犹豫、怀疑。那么能不能打破,这么一个怪圈呢?在沙漠植物里面它有很强大的根系,这些根系可以帮助植物获得充分的水分。然而植物根冠比,它作为一种应对干旱的弹性的策略,长久以来它的机制很难解决,为什么干旱它抑制植物地上部分的生长。但是促进根系的生长,为什么旱区的瓜果特别的甜?这些问题伴我从少年到中年。

在这个过程里面,我就首先问了一个问题,脱落酸是否起了作用?我们就做了脱落酸的多突变体,敲除了12个脱落酸的受体。最终发现在野生型里面干旱它可以抑制地上部分的生长,但是突变体里面,我们看不到这种抑制。野生型里面我们看到干旱促进根系的生长,突变体中没有这么一个响应,因此这就说明,干旱下植物利用脱落酸去调控根冠比。

下一个难题就来了,脱落酸如何调控呢?在普渡大学的一个学术讲座里面,我意外地发现,蔗糖转运突变体,它的表型和脱落酸的突变体非常相像,也是短根、植株矮小,种子有缺陷。当时就灵光一现,难道脱落酸通过调控蔗糖的转运,从而去影响植物根系的生长,植物整株的生长以及种子的灌浆,果实的甜度这些过程吗?其实科学很奇妙,我们有时候很多问题百思不得其解,但是有可能在特殊的契机下就会顿悟。当时我看到这个结果之后,我特别地兴奋,思维真的在那一瞬间就在奔逸。然后回到实验室马上设计实验,去检测我这个假设是否是这样的。然后当然,那位研究者后来成了我的合作者,我们一起去探讨了干旱胁迫下蔗糖转运,和植物根系生长的问题。
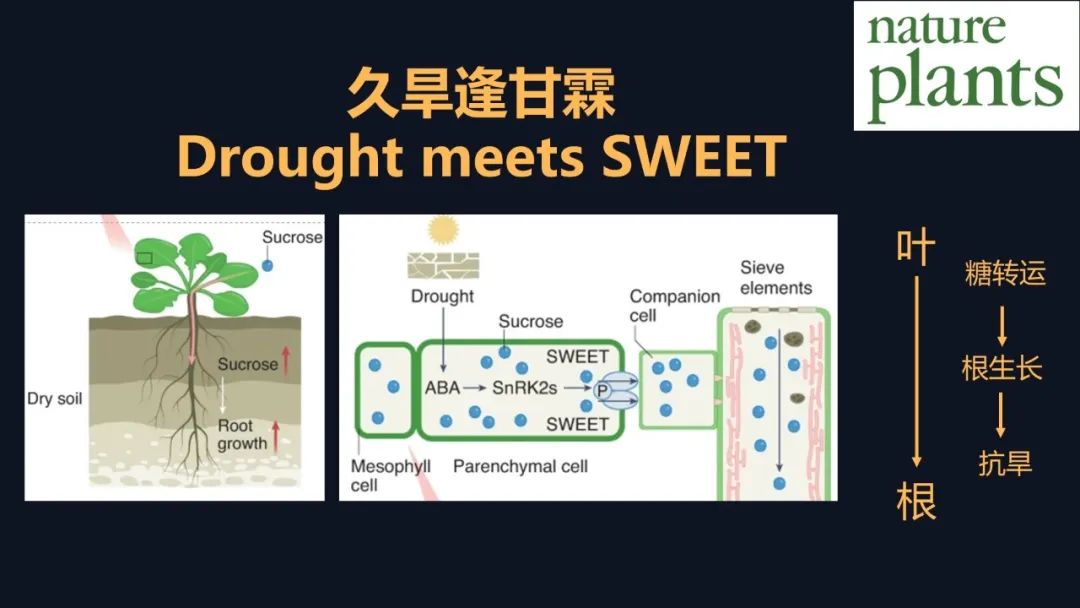
在经过五年的研究之后,我们才初步对这个机制得到了一些见解。干旱下由于根系的生长,它依赖于地上部分光合产物的转运,但是光合产物长距离的转运,它是以蔗糖的形式,蔗糖的转运它依赖于韧皮部,这就像一个高速路,而蔗糖转运蛋白SWEET,就像高速路的入口一样。干旱下脱落酸,它对蔗糖转运蛋白的调控提高了它的效率,我们发现了这么一个调控,它就像打通了植物体内,蔗糖的一个物流的转运,让叶片合成的这种蔗糖快速地转运到需要它的地方,就像根、芽和果实中,从整体上提高了植物的抗旱和生长。最终这个工作在年初发表在《自然-植物》上。同期还发表了中国农业大学,巩志忠教授和杨淑华教授,他们发表了评论《Drought meets SWEET》。这是很有一个寓意的名字,他们后来告诉我,其实是“久旱逢甘霖”。
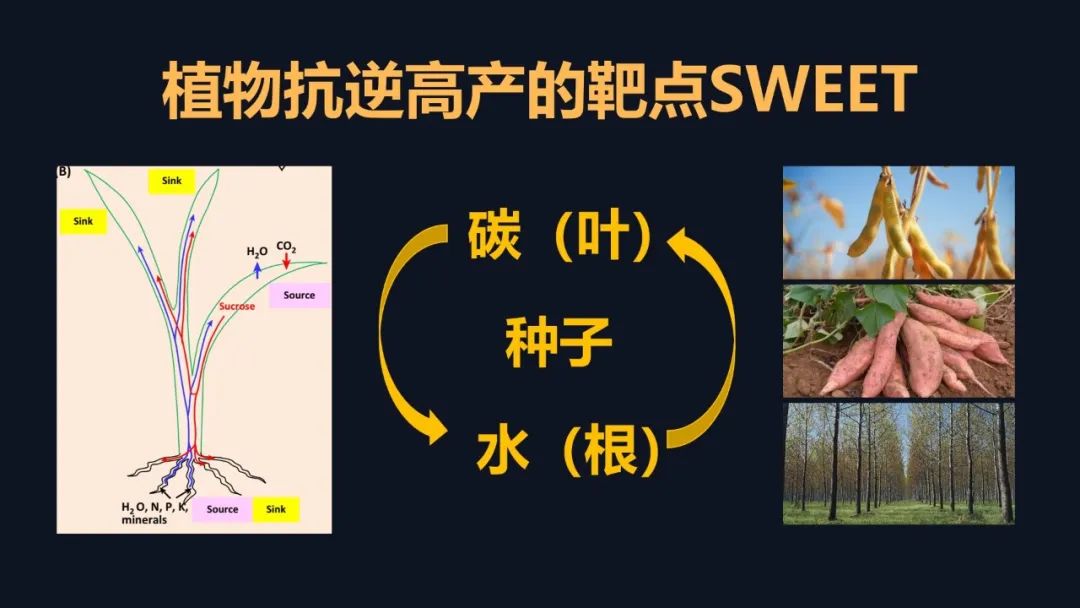
SWEET的调控,它还有上升空间吗?其实是有的。SWEET其实在进化上很奇特,它在进化上并不完美。它的活性非常的低,但是它的功能又非常的重要。大家想一想叶片上合成光合产物之后,需要把蔗糖快速地转运到需要它的地方,才能促进这些地方的生长,而根系生长了以后,才能提供充足的水分和养分,给我们植物整株的生长,所以如果我们进一步提升SWEET的活性,可以从一个开源的角度,提升植物整体的生长,抗逆和产量。当然它的应用空间非常的广泛,就像我们的种子植物、薯类,以及树木这些里面,我们的目标其实就是它进化不完美,我们能否通过我们的努力让它变得更完美。
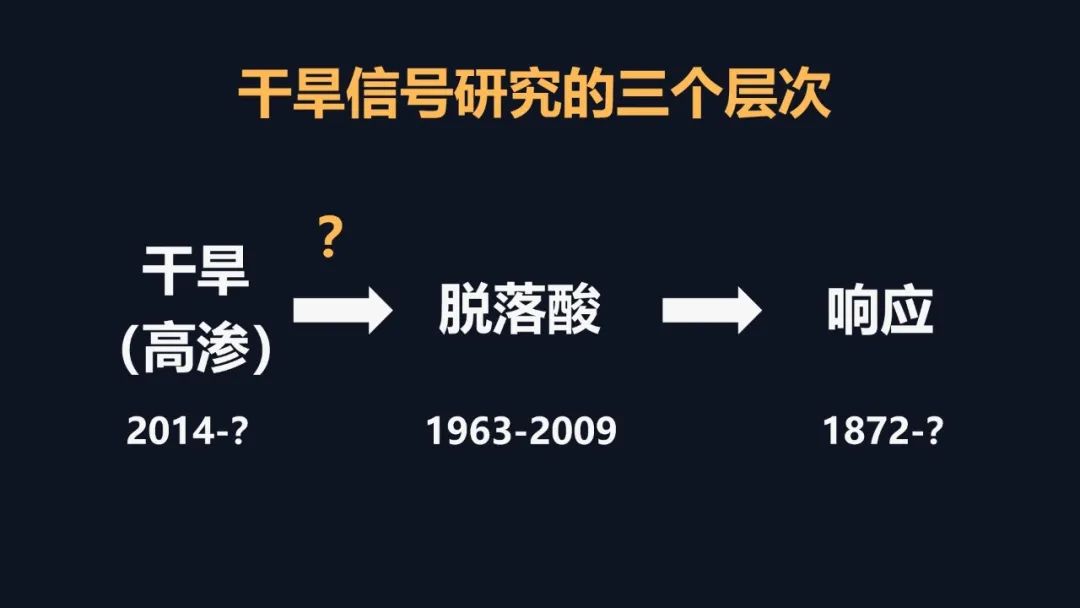
我们在前期的研究里面取得了一些进展,解决了一些干旱相关的问题,但是要把干旱相关的问题彻底解决,我们需要回到干旱的信号上,要去解决干旱信号的早期的感受和传导,但是早期的信号如何去研究呢?这里面我们有一个假设,我们把植物想象成一个军队,当它感知干旱之后,它有一个司令部可以做出来决策,发出命令,层层传递,从而调动整株植物,就像调动整个军队一样去抵御干旱。这个时候如果司令部成员出现了问题,那么决策不能做决策,命令不能传递就断掉了,但是如何去寻找呢?因为命令的传递和信号的传递它们都非常类似,你经过越多的环节,它越容易产生干扰,因此我们就用最早期的一个信号——钙信号去研究干旱的感知。
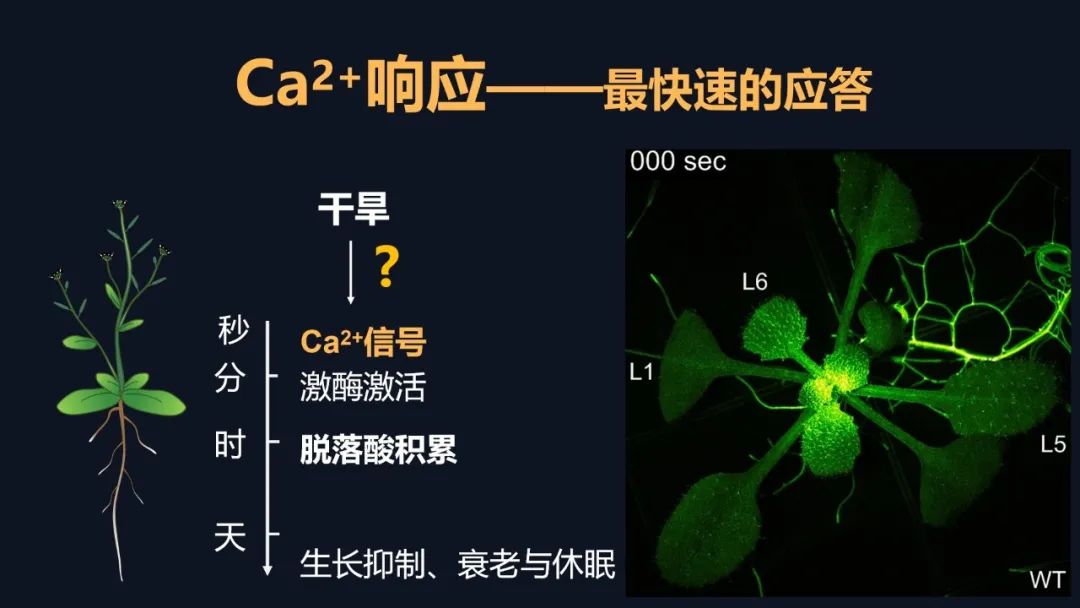
钙信号它是非常快的,但是有多快呢?大家看到了,剪掉一片叶子之后,钙信号瞬间爆发并且传遍植物的全身。植物它并不是不能感知,它只是不会跑而已。钙信号相关的研究,我们起始于多年前。大概在十七年前,因为我们要找这个司令部的成员的话,我们其实就是要找到钙信号减弱的突变体材料。但是十七年前,我在北京生命科学研究所还在读研,在我的老师的指导下,我们找到了钙信号增强的材料,但是没有找到钙信号减弱的材料。因为当时的钙信号非常的弱,找强的容易,找弱的难。因此在十年前,我做博后期间,我又用钙信号增强的材料建立群体,接着寻找钙信号弱的材料,并在两年后才找到第一株。当然这个材料的克隆就是我们平常的克隆等等,研究工作非常的艰难。这可能又过了六年之后,在我自己实验室又建立之后,四年才初步理解了,它感应干旱以及传递信号的潜在机制。这个工作才发表在《当代生物学》上。
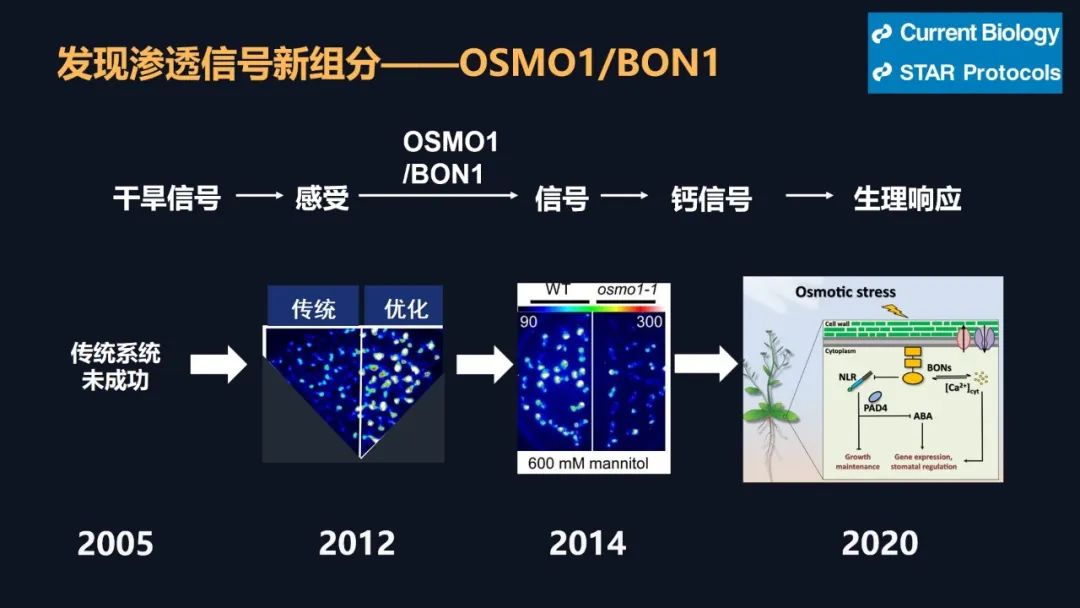
当然在这期间,我也是很长时间怀疑过自己,我的博士导师和我的博后导师,他们都跟我说过一个事情,他们说,我博后期间做这个事情,你肯定做不完,找工作的时候用不到。但是我想,有一些事情总要去做,总要去解决。他们都是非常支持我的。大家可以看到这个工作后面,有我的博士导师的支持,博后导师的支持,还有我实验室的成员的特别艰难的努力。当然我们现在实际上,只找到了司令部成员中的一员,我们还在尝试去找其它的几位,尤其是最关键的做决策的那几位。

在干旱的研究里面,有几个特别核心的问题。干旱是什么?干旱如何被植物感知?以及植物如何响应?我们如何做抗逆的作物?这些问题它都很难,但是也都能做,需要时间。“道阻且长,行则将至。”我们实验室就想着要逐步地解决,植物感应 传递,响应干旱的这些机制,并且把我们获得的知识,应用在作物的抗逆遗传改良上。我们期望获得这种抗逆稳产,高质量节水的农作物,然后让我们在干旱下至少还有吃的,让我们节约农业的用水,让我们在未来不至于过于恐慌。当然这非常艰难,但是“路漫漫其修远兮,与志同者共求索。”我们坚信有目标有同伴,我们可以解决这些问题。
谢谢大家!


| 留言与评论(共有 0 条评论) “” |